Survey Methods
Overview
Surveying for moths is most often and easily accomplished by setting lights at night to attract adults. Moths are primarily nocturnal, and most species are attracted to lights, some much more strongly than others. A single bright ultraviolet or incandescent light placed in a rich temperate habitat will reliably attract well over 100 species of moths on on calm, moonless, and warm summer nights. Even in quite degraded and fragmented semi-natural habitats, such as the post-agricultural meadows and shrublands that cover much of Block Island, a light can attract over 100 species of moths on the best nights. While certain moth taxa are diurnal or not attracted to lights, most species present in any given area will come to lights at least occasionally.
Light surveying has been the main method used in the Block Island moth survey, and only a handful of the more than 1,200 species known from Block Island have been recorded on the island only by other survey means. The most conspicuous and diverse group of Lepidoptera not attracted to lights is the butterflies, which comprise a single large diurnal superfamily, Papilionoidea. (Butterflies evolved from moths and from the perspective of cladistics are a group of diurnal moths in the same way birds are therapod [dinosaurs], hominids are apes, and ants are wasps.) Moths are also commonly surveyed by searching flowers on warm sunny days for nectaring diurnal adults, sweeping fields for adults, and searching for larvae on and in plants; I have not utilized these methods as much as I would like so far.
What makes my survey nearly unique among moth surveys is that since 2018, I have made comprehensive records of abundances of all species each survey night, recording more than 40,000 individual moths in four seasons. This dataset allows for quantitative analysis of the fauna, which I have only just begun. For each species, I have generated a flight time phenology chart and a map of relative abundance in each area surveyed. For common species, my observations reliably measure flight times and habitat preferences. My data can also be used to characterize Block Island’s moth fauna in great detail. So far, I have produced measures of the similarity of the fauna between survey locations and, using my observations at my porch lights, its change over the course of the season.
Timeline
The methods, goals, and scope of this survey evolved organically over the course of several years beginning in 2014. This survey began as part of a more general survey of Block Island’s animal fauna I began in August 2014 with no guidance or prior experience in faunal surveying methods or species identification. Over the following few years, moths become nearly the exclusive focus of the project, and the moth component of my survey gradually grew into an effort at a comprehensive survey of Block Island’s moths. I began collecting moths in 2016 but did not intensively collect for identification until 2018. I began blacklighting around the island, collecting smaller microlepidoptera, and counting common species at my porch lights in 2017. In 2018, my survey efforts took close to their definitive form. I regularly set up sheets for blacklighting at sites across the island that summer, recorded all individual moths sighted both at my porch lights at at blacklighting sheets, and intensively collected taxa I could not identify to species by sight. In 2021, I began using no-kill bucket traps to supplement my surveying with sheets, particularly in dunes, where winds slacken enough for surveying with sheets only a few nights each summer. Future plans, aside from additional surveying for moths, include floristic and habitat surveys of each location sampled and a detailed plant inventory of the Hunt property and surrounding area.
I began nightly estimates of species counts at my porch lights in 2017 for a set slate of several dozen common species and species complexes and in 2018 for nearly every species. Certain groups, such as Pero spp. and Eulithis spp., in which the species present on Block Island are difficult to impossible to separate on sight, I have recorded jointly. Some groups I could not personally identify on sight in 2018 or into 2019, notably all Renia spp., and some Acronicta spp., I always photographed and/or collected for later determination. Anything unrecognized I likewise photographed and/or collected. Since 2019, I have always collected each species I have not previously seen. For Coleophora spp. and Blastobasidae, I recorded morphospecies as precisely as I could, though I shortsightedly did not consistently make note of the streaky tan Coleophora spp. in 2019. I have since identified many of these morphospecies in through identification of collected specimens, though probably several of Block Island’s Coleophora species are indistinguishable by sight due their similarity, low contrast patterning, and small size. Aside from the omission of some Coleophora spp. in 2019, I have recorded in some manner all moths seen since 2018.
While the great majority of my survey effort has been with lights, I have searched in daytime for adults of diurnal species on flowers and vegetation and by sweeping meadows and for larvae, especially leafminers. Additionally, Nigel Grindley has regularly photographed and collected moths at lights at his home in the southwest of Block Island for several years, by a remarkable coincidence starting the same year I did. In the last few years, he has also surveyed at sheets with blacklights several years while I have been off the island. By my count, he has found 58 species I have not found, of these, fewer than ten were recorded by previous surveyors, making Nigel’s contribution to the island’s species list quite significant. (He has of course seen several hundred of the species I have seen as well.) His surveying has disproportionately added species that fly only very early or late in the season, when I have rarely been on Block Island.
Recording Sightings
The methodology I have used for counting numbers of each species at my porch lights is roughly as follows: I start by scanning the porch, walls, and eaves for moths and note the species I see on my phone. As I go, I photograph uncommon or otherwise interesting moths and capture specimens I wish to collect, usually for identification. I frequently update estimated numbers of each species based either on spotting specimens that were worn or differently patterned than those previously counted or by scanning the entire area for all examples of a set of species for a more complete count. For especially abundant species, I estimate numbers. While my methodology is highly inexact, especially during the most productive nights, it has been fairly consistent. The time I spend at the porch lights each night varies based on the number of moths present, but during the peak of the season, 45-60 minutes is probably typical.
During blacklighting sessions, I follow a similar procedure to that I employ at my porch lights, except in that I spend most or all of the night observing moths until I take down the sheets. When I use two sheets, as I typically do, I record moths separarately at each sheet. I make multiple visits to each sheet each night, and I often note the moths I see flying past when walking between the sheets. My notes on moth found away from the sheets have informed my understanding of which common species are poorly drawn to lights. My typical procedure for counting individuals of a species at a sheet is to quickly but thoroughly scan each side of the sheet and then any nearby plants as well as the ground, continually scanning the sheet and updating counts as long as the total number of individuals present appears to be increasing. As at the porch lights, I use photography and collection to fill all gaps in the data created by identification uncertainty, excepting certain species complexes.
Data Analysis
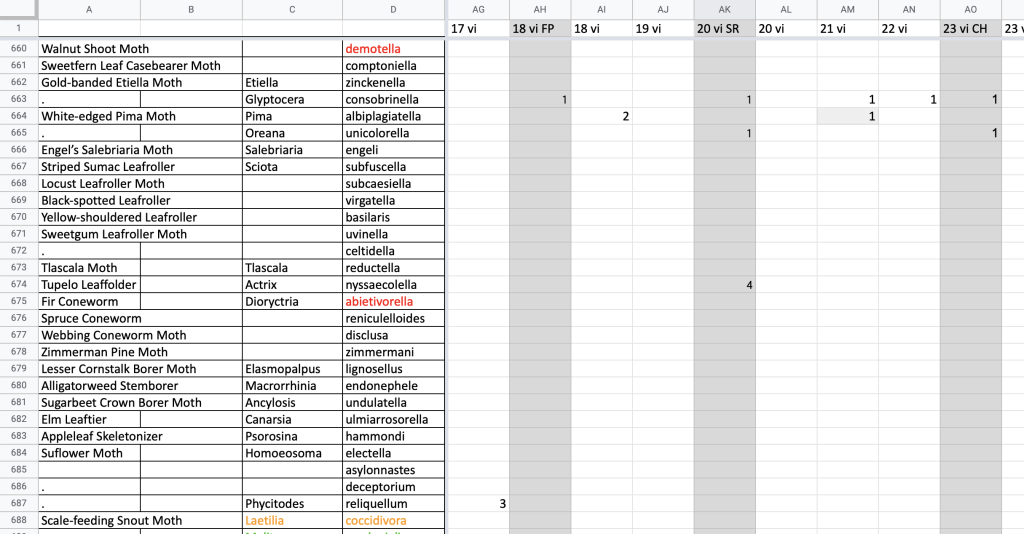
At the end of each season, I compile all my sighting records into a single document in chronological order. I manually add my sightings to a spreadsheet with a line for each species and a column for each night or blacklighting sheet (Fig. 1). Records of unidentified species are marked in red and with asterisks and revisited when I identify the species they represent. I add photographic and specimen records, not all of which I include in my nightly notes, to my tables to complete my records.
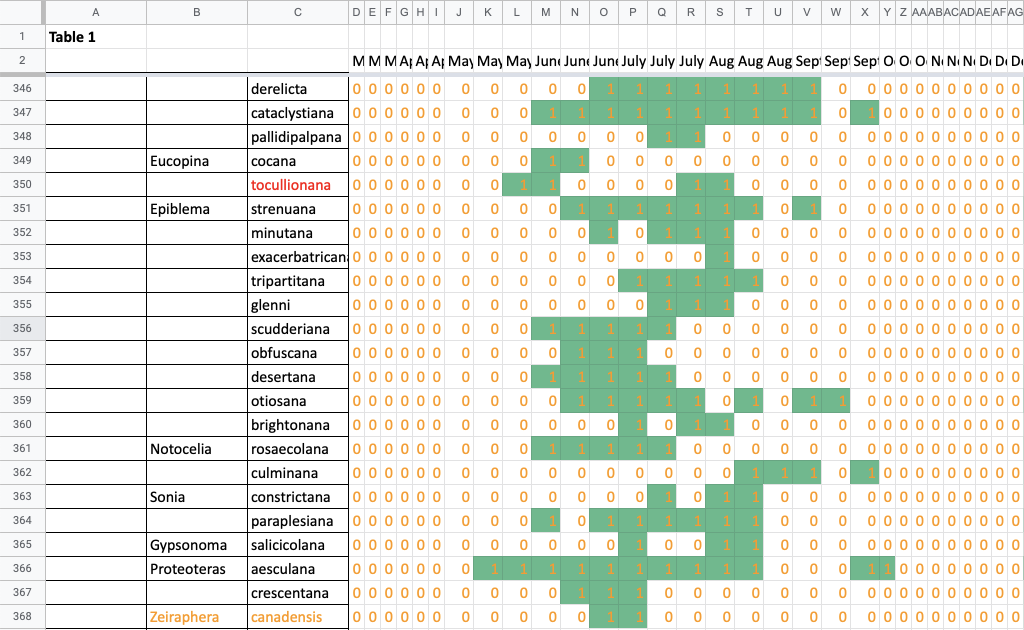
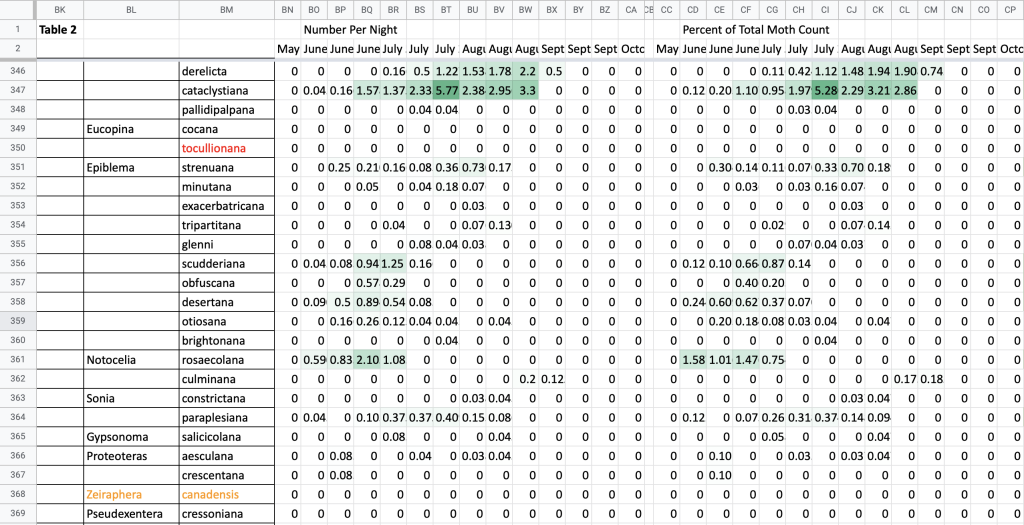
I use a series of spreadsheets to store all my photographic records from 2017 and earlier, Nigel’s photographic records, and records from older surveys; and I use these along with my 2018–2021 sighting tables to make a single table with presence/absence data for each species in each 1/3-month period from March through December (Fig. 2). I use the sighting records from my porch lights from 2018 to present to calculate flight time phenology in individuals per survey night and as a percent of all moth records for each 1/3-month period (Fig. 3).
Partial List of Relevant Survey Data and Other Research
*Not open access
^Based on systematic (Rothamsted Insect Survey and Garden Moth Scheme) and opportunistic (National Moth Recording Scheme) population monitoring in British Isles
Major North American Moth Phenology Data Sets
GA: Clarke Co. (J. Pickering) — Hundreds of thousands of photographic records since 2010. This is the only other complete all-taxa nightly moth abundance data set of which I am aware. Sighting data on individual species can be found here.
NC (Moths of North Carolina) — >150,000 records of moths in the state, compiled by S. Hall et al. from all available sources (collected specimens, survey reports, and photographs). Adult phenology charts for each species are generated separately for each of four ecoregions: high mountains, low mountains, piedmont, and coastal plain.
MA (MassMoths) — >250,000 records of moths in the state, compiled by S. Whitebread et al. from all available sources (collected specimens, survey reports, and photographs). Currently, adult (and larval) phenology charts are shown for only a small number of species, e.g. 1, 2, 3.
OR, WA, BC (PNWMoths) — Nearly 100,000 records, mostly museum specimens, of 1,248 species of macromoths (Macroheterocera) excluding Geometridae. Most records are from Oregon, Washington, and southern British Columbia; Idaho and adjacent portions of surrounding states are also covered. Phenology is graphed for each species, and records can be filtered by elevation and year.
LA: Abita Springs (V.A. Brou) — Millions of records of moths from large-scale nightly light-trapping, conducted continuously since 1969. Several dozen published short papers, each treating one species or a small genus, some coauthored by C.D. Brou, report summarized phenology data along with state distribution data. These can be found in the complete archives of the Southern Lepidopterists’ News, which is indexed for easy searching.
Phenology, Population Variation, and Local Biogeography
Albu, V. & S. Albu, 2023. Lepidoptera assemblages along a western slope elevation gradient of the south-central Sierra Nevada mountains in California. J. Lep. Soc. 77(1): 43–58.*
Highland, S.A., J.C. Miller, & J.A. Jones, 2013. Determinants of moth diversity and community in a temperate mountain landscape: vegetation, topography, and seasonality. Ecosphere 4(10): 129.
Mustelin, T., L.G. Crabo, & C. Looney, 2020. Lepidoptera Diversity Based on Continuous Surveys 2009–2011 and 2017–2019 at a Transition Zone Site in the Central Cascade Range, Washington, USA. J. Lep. Soc. 74(4): 263–289.*
Powell, J.A., 1994. Biogeography of Lepidoptera on the California Channel Islands, pp. 450–464 in W.L. Halvorson & G. J. Maender (eds.). The Fourth California Islands Symposium: Update on the Status of Resources. Santa Barbara Museum of Natural History, Santa Barbara, CA.
Raimondo, S., A.M. Liebhold, J.S. Strazanac, & L. Butler, 2004. Population synchrony within and among Lepidoptera species in relation to weather, phylogeny, and larval phenology. Ecological Entomology 29(1): 96–105. [PDF]
Stange, E.E., M.P. Ayres, & J.A. Bess, 2011. Concordant population dynamics of Lepidoptera herbivores in a forest ecosystem. Ecography 34: 772–779.
Wilson, J.F., D. Baker, M. Cook, G. Davis, R. Freestone, D. Gardner, D. Grundy, N. Lowe, S. Orridge, & H. Young, 2015. Climate association with fluctuation in annual abundance of fifty widely distributed moths in England and Wales: a citizen-science study. J. Insect Conserv. 19: 935–946.^
Work, T.T. & D.G. McCullough, 2000. Lepidopteran Communities in Two Forest Ecosystems During the First Gypsy Moth Outbreaks in Northern Michigan. Environ. Entomol. 29(5): 884–900.
Other North American Phenology Data
Lewis, M.N., R.M. Steichen, & K.S. Summerville, 2005. The Moths of Neal Smith National Wildlife Refuge: A Preliminary Assessment. Journal of the Iowa Academy of Science: JIAS 112(1–2): 1–12.
Murphy, S.M., J.T. Lill, & M.E. Epstein, 2011. Natural history of limacodid moths (Zygaenoidea) in the environs of Washington, D.C. J. Lep. Soc. 65(3): 137–152.
Tolley, M.P., 1982. Aspects of the biology of sod webworms (Lepidoptera: Pyralidae: Crambinae) and turfgrass inhabiting insects of Virginia. MS entomology thesis at Virginia Tech. 52 pp.
Forest Defoliator Population Cycling
Büntgen, U., A. Liebhold, D. Nievergelt, B. Wermelinger, A. Roques, F. Reinig, P.J. Krusic, A. Piermattei, S. Egli, P. Cherubini, & J. Esper, 2020. Return of the moth: rethinking the effect of climate on insect outbreaks. Oecologia 192: 543–552.
Haynes, K.J., A.J. Allstadt, D. Klimetzek, 2014. Forest defoliator outbreaks under climate change: effects on the frequency and severity of outbreaks of five pine insect pests. Global Change Biology 20(6): 2004–2018.*
Nealis, V.G. & J. Régnière, 2014. An individual-based phenology model for western spruce budworm (Lepidoptera: Tortricidae). The Canadian Entomologist 146(3): 306–320. [PDF]
Urban Moth Communities
Aagaard, K., K. Berggren, P.D.N. Hebert, J. Sones, B. McClenaghan, & T. Ekrem, 2017. Investigating suburban micromoth diversity using DNA barcoding of malaise trap samples. Urban Ecosyst. 20: 353–361.
Altermatt, F. & D. Ebert, 2016. Reduced flight-to-light behaviour of moth populations exposed to long-term urban light pollution. Biol. Lett. 12: 20160111.
Bates, A.J., J.P. Sadler, D. Grundy, N. Lowe, G. Davis et al., 2014. Garden and landscape-scale correlates of moths of differing conservation status: significant effects of urbanization and habitat diversity. PLoS One 9(1): e86925.^
Boyes, D.H., D.M. Evans, R. Fox, M.S. Parsons, & M.J.O. Pocock, 2020. Is light pollution driving moth population declines? A review of causal mechanisms across the life cycle. Insect Conserv. Divers. 14(2): 167–187.
Ellis, E.E. & T.L. Wilkinson, 2021. Moth assemblages within urban domestic gardens respond positively to habitat complexity, but only at a scale that extends beyond the garden boundary. Urban Ecosystems 24: 469–479.
Franzén M., P.-E. Betzholtz, L.B. Pettersson, & A. Forsman, 2020. Urban moth communities suggest that life in the city favours thermophilic multi-dimensional generalists. Proc. R. Soc. B 287: 20193014.*
Kadlec, T., M.A.A.M. Kotela, I. Novák, M. Konvička, & V. Jarošík, 2009. Effect of land use and climate on the diversity of moth guilds with different habitat specialization. Community Ecology 10(2): 152–158.
Lintott, P.R., N. Bunnefeld, E. Fuentes-Montemayor, et al., 2014. Moth species richness, abundance and diversity in fragmented urban woodlands: implications for conservation and management strategies. Biodivers. Conserv. 23: 2875–2901.
Merckx, T. & H. Van Dyck, 2019. Urbanization-driven homogenization is more pronounced and happens at wider spatial scales in nocturnal and mobile flying insects. Glob. Ecol. Biogeogr. 28(10): 1440–1455.*
Plummer, K.E., J.D. Hale, M.J. O’Callaghan, J.P. Sadler, & G.M. Siriwardena, 2016. Investigating the impact of street lighting changes on garden moth communities. J. Urban Ecol. 2(1): juw004.
Rice, A.J. & P.J.T. White, 2015. Community Patterns in Urban Moth Assemblages. J. Lep. Soc. 69(3): 149–156.*
Scalercio, S., P. Brandmayr, N. Iannotta, R. Petacchi, & L. Baccaccio, 2012. Correlations between landscape attributes and ecological traits of Lepidoptera communities in olive groves. Eur. J. Entomol. 109: 207–216.
Slade, E.M., T. Merckx, T. Riutta, D.P. Bebber, D. Redhead, P. Riordan, & D.W. Macdonald, 2013. Life‐history traits and landscape characteristics predict macro‐moth responses to forest fragmentation. Ecology 94(7): 1519–1530.*
Long-Term Changes
Blumgart, D., M.S. Botham, R. Menéndez, & J.R. Bell, 2022. Moth declines are most severe in broadleaf woodlands despite a net gain in habitat availability. Insect Conserv. Divers. 15(5): 496–509.^
Boyes, D.H., R. Fox, C.R. Shortall, & R.J. Whittaker, 2019. Bucking the trend: The diversity of Anthropocene ‘winners’ among British moths. Front. Biogeogr. 11(3): e43862.^
Burner, R.C., V. Selås, S. Kobro, R.M. Jacobsen, & A. Sverdrup‐Thygeson, 2021. Moth species richness and diversity decline in a 30‐year time series in Norway, irrespective of species’ latitudinal range extent and habitat. J. Insect Conserv. 25: 887–896.
Conrad, K.F., M.S. Warren, R. Fox, M.S. Parsons, I.P. Woiwod, 2006. Rapid declines of common, widespread British moths provide evidence of an insect biodiversity crisis. Biol. Conserv. 132(3): 279–291. [PDF]^
Dennis, E.B., T.M. Brereton, B.J.T. Morgan, R. Fox, C.R. Shortall, T. Prescott, & S. Foster, 2019. Trends and indicators for quantifying moth abundance and occupancy in Scotland. J. Insect Conserv. 23: 369–380.^
Fox, R., 2013. The decline of moths in Great Britain: a review of possible causes. Insect Conserv. Divers. 6(1): 5–19.^
Fox, R., E.B. Dennis, C.A. Harrower, D. Blumgart, J.R. Bell, P. Cook, A.M. Davis, et al., 2021. The State of Britain’s Larger Moths 2021. Butterfly Conservation, Rothamsted Research, and UK Centre for Ecology & Hydrology, Wareham, Dorset, UK. 44 pp.^
Fox, R., T.H. Oliver, C. Harrower, M.S. Parsons, C.D. Thomas, & D.B. Roy, 2014. Long-term changes to the frequency of occurrence of British moths are consistent with opposing and synergistic effects of climate and land-use changes. J. Appl. Ecol. 51(4): 949–957.^
Laussmann, T., A. Dahl, & A. Radtke, 2021. Lost and found: 160 years of Lepidoptera observations in Wuppertal (Germany). J. Insect Conserv. 25: 273–285.
Wagner, D.L., R. Fox, D.M. Salcido, & L.A. Dyer, 2021. A window to the world of global insect declines: Moth biodiversity trends are complex and heterogeneous. PNAS 118(2): e2002549117.
Wölfling, M., B. Uhl, & K. Fiedler, 2019. Multi-decadal surveys in a Mediterranean forest reserve – do succession and isolation drive moth species richness? Nature Conserv. 35: 25–40.
Caterpillar Surveys
Abbott, I., T. Burbidge, M. Williams, & P. Van Heurck, 1992. Arthropod fauna of jarrah (Eucalyptus marginata) foliage in Mediterranean forest of Western Australia: Spatial and temporal variation in abundance, biomass, guild structure and species composition. Aus. J. Ecol. 17(3): 263–274.*
Loeffler, C.C., 1992. Comparative structure of the lepidopteran faunae associated with goldenrods (Solidago spp.) in forests and fields. PhD entomology dissertation at Cornell University. 292 pp.*
Seifert, C.L., L.R. Jorge, M. Volf, D.L. Wagner, G.P.A. Lamarre, S.E. Miller, E. Gonzalez-Akre, K.J. Anderson-Teixeira, & V. Novotný, 2021. Seasonality affects specialisation of a temperate forest herbivore community. Oikos 130(9): 1450–1461.*
Pollinators
Wonderlin, N.E., K. Rumfelt, & P.J.T. White, 2019. Associations between Nocturnal Moths and Flowers in Urban Gardens: Evidence from Pollen on Moths. J. Lep. Soc. 73(3): 173–176.*
Other
Chapman, J.W., J.R. Bell, L.E. Burgin, D.R. Reynolds, L.B. Pettersson, J.K. Hill, M.B. Bonsall, & J.A. Thomas, 2012. Seasonal migration to high latitudes results in major reproductive benefits in an insect. PNAS 109(37): 14924–14929.
Forister, M.L., C.S. Philbin, Z.H. Marion, C.A. Buerkle, C.D. Dodson, J.A. Fordyce, G.W. Forister, S.L. Lebeis, L.K. Lucas, C.C. Nice, & Z. Gompert, 2020. Predicting patch occupancy reveals the complexity of host range expansion. Science Advances 6(48): eabc6852.